2021/04/26
(株)エコタイプ次世代植物工場 竹葉 剛
光合成といえば空気中の二酸化炭素を光のエネルギーを使って有機物に変換する反応系であると理解している方も多いと思う。これは炭素からみた反応系であり、窒素についてみると窒素同化、すなわち硝酸還元系が光合成の主要な反応系である。この他に、イオウの同化系もあり、整理すると、光合成とは、炭素(C)、窒素(N)、イオウ(S)の同化系から成り立っていることになる。いずれも、無機化合物から光のエネルギーを使って同化、すなわち、有機物を合成する反応系である。
窒素の同化系は、硝酸還元酵素(NR, Nitrate Reductase)、亜硝酸還元酵素(NiR, Nitrite Reductase)、グルタミン合成酵素(GS, Glutamine Synthetase)から構成されている。このうち、NRは細胞質に、NiR、GS は葉緑体内に分布している。GS は細胞質にも存在する。GSによりアミノ酸に取り込まれたグルタミンのアミノ基は、グルタミン酸や他のアミノ酸の合成に利用される。
種子発芽期には、根や芽が発育するので、多くのタンパク質が合成され、そのためのアミノ酸が多く必要となる。種子中には貯蔵タンパク質が含まれており、吸水とともにその貯蔵タンパク質の分解が始まる。生体のタンパク質を構成するアミノ酸は20種あるが、貯蔵タンパク質の分解によって生成した20種のアミノ酸がそのまま種子発芽に必要なタンパク質に利用されるのではなく、貯蔵タンパク質の分解で生成した20種のアミノ酸に含まれるアミノ基は一旦はすべてアンモニアにまで分解され、そのアンモニアはグルタミンに取り込まれて、種子発芽に必要なタンパク質合成の場である根や芽に移動して、そこでグルタミン酸や他のアミノ酸へ変換され、新たなタンパク質合成に利用される。なぜこのような複雑な過程を繰り返しているのかの理由は、結局のところ、必要なアミノ酸を必要に応じて供給するという合理的なしくみがある、と理解すべきである。
そのようにして、貯蔵タンパク質が窒素源として存在している間は、種子発芽の発育を支えている窒素源は貯蔵タンパク質である。貯蔵タンパク質が分解され尽くされるとその窒素源はなくなることになる。外部から観察してみると、種子が吸水し、子葉が展葉し始めるころまでは、貯蔵タンパク質が窒素源として機能している、とみなしてよい。もちろん、貯蔵タンパク質の量は種子の種類によって大きく異なるので、貯蔵タンパク質が種子発芽の発育を支えている時期は同じではない。
貯蔵タンパク質がなくなった後に窒素源となるのは、種子外部から吸収した硝酸イオンである。硝酸イオンがNR, NiR, GS の働きでアンモニア、グルタミンに取り込まれる硝酸還元系が窒素供給の重要な担い手になる。このように考えれば、種子発芽期の初期から硝酸イオン(塩)を供給しておけば、窒素源の供給がスムーズに進行すると考えられる。しかし、種子発芽の初期から硝酸イオンを与えておけば、根の伸長が強く抑制され、その後の発芽過程がかえって遅れる、という現象が起きる。この原因はまだよく分かっていないが、硝酸還元系の作動には多くのエネルギーが必要となるので、発芽初期から硝酸還元系を作動させると、エネルギーがそのために消費され、他の反応系へのエネルギー供給が抑制されるためではないか、と考えられる。そのため、植物工場で苗をつくる場合には、成育期の養液を希釈してEC値を半分程度に薄めて育苗することが経験的に行われている。
ところが、成育期の養液を希釈して育苗する方法には、問題点が二つある。一つは、希釈によってマグネシウムなどの濃度が低下して、発育が抑制されることである。成育期の養液(大塚A処方)をEC=1.8から1/2に希釈すると、Mg 濃度は約12 ppmとなる。MgはDNA polymerase活性の維持に必要であるから、希釈によって成長点での細胞分裂に支障が生ずる。問題点の第二は、育苗期間を2週間とすれば、育苗期間の間、苗の発育が相対的に抑制された状態になり、そのような苗を移植しても収穫までの生育が相対的に遅れることなる点、である。第二の問題点は、成育期の養液を希釈すると、硝酸イオンの濃度が低くなり、そのため硝酸還元系の誘導が不十分になる、ためではないかと考えられる。
上記の問題点を解決するために、発芽初期に根が硝酸イオンによって抑制される期間を調べてみると、わずか2~3日間であることが判明した。そのため、レタス、コマツナなどの通常の野菜種子の場合、まず水で4日間発芽させ、その後2~3日間はMg, Zn などのミネラル成分濃度を上げて、硝酸イオン濃度を抑えた第一循環養液で養液供給を行い、その後で第二循環養液として、硝酸イオン、Mg, Zn濃度を高めた養液を供給すれば問題は解決する。ここで、硝酸還元系酵素(Nr, NiR)はいずれも誘導酵素であり、比較的高濃度の硝酸イオン(数mM 以上)が必要とされる。また、アンモニアは誘導を抑制するので、大塚液肥でい、1号、5号、硫安などは使用しない方がよい。
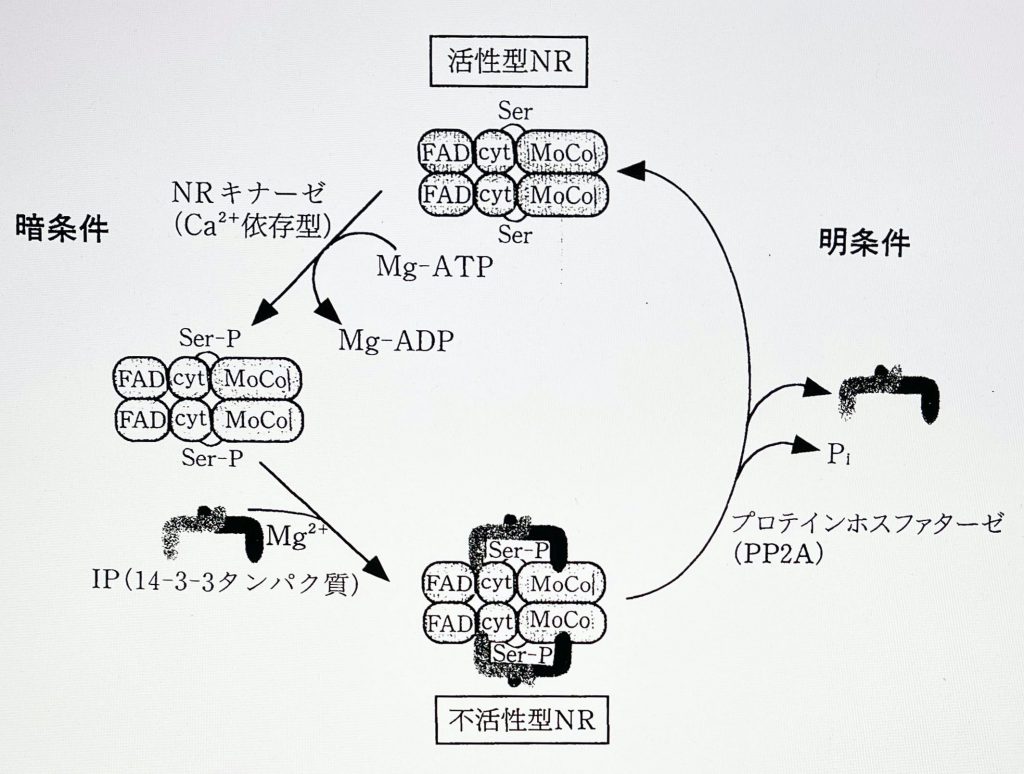
ところで、硝酸還元系は明暗条件下で活性調節も受けており、明で活性化、暗で不活性化する(図は、活性型NRと不活性型NR とがCa++, Mg++ により調節を受けることを説明;化学と生物 vol 36, 1996)。その調節は、暗条件下で NR が細胞質のCa++ 、Mg++ の濃度上昇によって不活性化され、明条件下で葉緑体内にCa++, Mg++ が取り込まれて細胞質のCa++, Mg++濃度が低下することによって活性化される、という調節を受けている。これは、暗条件下ではエネルギーの供給がなくなるので、NR活性を阻害することでエネルギーバランスを保っているため、と考えられている。この際に問題となるのが、Ca++, Mg++ 濃度である。特に、Mg++濃度を高くしてDNA Polymerase 活性を高く保とうとすると、細胞質のCa++, Mg++ 濃度が明条件下でも高くなり、明条件下でNR活性が抑制され、硝酸還元系が阻害される結果となり、葉は次第に黄化してくる(黄化はアブラナ科のコマツナで顕著であり、レタス類ではあまり顕著ではない)。明条件下でNR活性を維持するためには、Mg++濃度を上げすぎないことが肝心である。一応の目安として、第二循環養液にはA処方(S1)のみを用いると、黄化は起きない。
以上を要約すると、種子発芽期では、まず根の伸長を保証するために4日間の吸水の後、2~3日間硝酸イオン濃度を下げ、Mg++, Zn++濃度を上げた第一養液で循環させ、次いで硝酸イオン濃度を十分上げて硝酸還元系を誘導し、その後NR活性を維持するために適度なCa++ 、Mg++ 濃度を保つことが、丈夫な苗を育成し、ひいては収穫時期を早める栽培方法であるといえる。